Alfredson H, Pietilä T, Jonsson P and Ronny L (1998): Heavy-Load Eccentric Calf Muscle Training For the Treatment of Chronic Achilles Tendinosis. The American Journal of Sports Medicine 26: 360-366.
Alfredson H and Lorentzon R (2000): Chronic Achilles Tendinosis Recommendations for treatment and Prevention. Sports Medicine 29: 135-146.
Archambault DM, Wiley JP and Bray RC (1995): Exercise loading to tendon and the development of overuse injuries, a review of current literature. Sports Medicine 20: 77-89.
Binkley J (1989): Overview of ligament and tendon structure and mechanics: implications for clinical practice. Physiotherapy Canada 41: 24 – 31.
Boorman RS, Shrieve NG and Frank CB (1998): Immbolization increases the vulnerability of rabbit medial collateral ligament autografts to creep. Journal of Orthopaedic Research 16: 682 – 689
Burgeson RE and Nimni ME (1992): Collagen Types. Molecular Structure and Tissue Distribution. Clinical Orthopaedics and Related Research 282: 250-273.
Culav EM, Clark CH and Merrilees MJ (1999): Connective tissue: Matrix composition and its relevance to Physical Therapy. Physical Therapy 79: 308-319.
Frank C, Shrive N, Hiraoka H, Nakamura N, Kaneda Y and Hart D (1999): Optimisation
of the biology of soft tissue repair. Journal of Science and Medicine in Sport 2: 190 – 210.
Frank CB (1996): Ligament healing: current knowledge and clinical applications. Journal
of American Academy of Orthopaedic Surgeons 4: 74 – 83.
Frankel V and Nordin M (1980): Basic biomechanics of the skeletal system. Philadelphia: Lea & Febiger.
Gray’s Anatomy (38th ed.) (1995): London: Churchill Livingstone.
Gum L S, and Reddy G K (1997): Combined Ultrasound, Electrical Stimulation, and Laser Promote Collagen Synthesis with Moderate Changes in Tendon Biomechanics. American Journal of Physical Medicine & Rehabilitation 76: 288-296
Hall BK, Miyake T (1992): The Membranous skeleton: the role of cell condensations in vertebrate skeletogenesis. Anatomy and Embryology. Review, Tutorial 186(2):107-24.
Hayman J and Rodeo SA (2000): Injury and repair of tendons and ligaments. Physical
Medicine and Rehabilitation Clinics of North America 11: 267 – 288.
Hildebrand KA and Frank CB (1998): Scar formation and ligament healing. Canadian
Journal of Surgery 41: 425 – 429.
Hukins D W L (Ed) (1990): Connective Tissue Matrix (Part Two). Hampshire: The Macmillan Press.
Hukins DW (1990): Dynamic Aspects of Connective Tissue Structure and Function. London: Macmillan Press.
Huijbregts PA and Smith SE (1999): Tendon Injury: A Review. The Journal of Manual
and Manipulative Therapy 7: 71-80.
Hunter G (1994): Specific Soft Tissue Mobilisation in the Treatment of Soft tissue Lesions. Physiotherapy 80: 15-20.
Hyman J and Rodeo SA (2000): Injury and Repair of Tendons and Ligaments. Physical Medicine and Rehabilitation Clinics of North America 11: 267-288.
Jozsa L and Kannus P (1997): Human Tendons: anatomy,physiology and pathology: Human Kinetics.
Kennedy J, Hawkins R, Willis R and Danylchuck K (1976): Tension studies of human knee ligaments. Yield point, ultimate failure and disruption of the cruciate and tibial collateral ligaments. The Journal of Bone and Joint Surgery 58: 350-355.
Leadbetter WB (1992): Cell-Matrix Response in Tendon Injury. Clinics in Sports
Medicine volume 11: 533 – 578.
Liu SH, Yang R-S, Al-Shaikh R and Lane JM (1995): Collagen in tendon, ligament, and
bone healing: a current review. Clinical Orthopaedics and Related Research 318: 265 –
278.
Luciano, Dorothy S.(1978): Human Function and Structure: New York: McGraw-Hill, Inc.
Mayne R (1997): Arthritis and Allied Conditions. A Textbook of Reumatology (13th ed). Baltimore: Williams & Wilkins.
Nigg B and Herzog W (1999): Biomechanics of the musculoskeletal system. (2nd ed.) New York: John Wiley & Sons.
Noyes F (1977): Functional properties of knee ligaments and alterations induced by immobilisation. Clinical Orthopaedics 123: 210-242.
Noyes F and Grood E (1976): The strength of the anterior cruciate ligament in humans and rhesus monkeys. The Journal of Bone and Joint Surgery 58: 1074-1082.
Noyes F, Torvik P and Hyde W (1974): Biomechanics of ligament II. An analysis of immobilisation, exercise and reconditioning effects in primates. The Journal of Bone and Joint Surgery 56:
O’Brien M (1992): Functional anatomy and physiology of tendons. Clinics in Sports
Medicine 11: 505-520.
Palastanga N, Field D and Soames R (1994): Anatomy and Human Movement, Structure And Function (2nd ed.) Oxford: Butterworth Heinemann
Panjabi M and White IA (2001): Biomechanics in the musculoskeletal system. New York: Churchill Livingstone.
Reid D (1992): Sports injury assessment and rehabilitation. New York: Churchill Livingstone.
Reid DC (1992): Sports Injury Assessment and Rehabilitation. New York: Churchill & Livingstone.
Rogers, Andrew W. (1983): An Introduction to Histology and Cell Biology: London: Academic Press Inc.
Rundgren A (1974): Physical properties of connective tissue as influenced by single and repeated pregnancies in the rat. Acta Physiol Scandanavia Suppl: 417.
Sandrey MA (2000): Effects of acute and chronic pathomechanics on the normal
histology and biomechanics of tendons:a review. Journal of Sport Rehabilitation 9: 339-
352.
Schultz R, Miller D, Kerr C and Micheli L (1984): Mechanoreceptors in human cruciate ligaments. The Journal of Bone and Joint Surgery 66A (7): 1072-1076.
Watkins James, (1999): Structure and Function of the Musculoskeletal System.
Williams CJ, Vandenberg P and Prockop DJ (1997): Textbook of Rheumatology (5th ed). Philadelphia: W.B. Saunders Company.
Williams PL, Bannister LH, Berry MM, Collins P, Dyson M, Dussek JE and MWJ Ferguson (Eds) (1995). Gray’s Anatomy (38th ed). New York, Churchill Livingston: 75-90.
Wilson AM and Goodship AE (1994): Exercise-induced hyperthermia as a possible mechanism of tendon degeneration. Journal of Biomechanics 27: 899-905.
Woo S, Gomez M and Inoue M (1983): Measurement of mechanical properties of ligament substance from a bone-ligament-bone preparation. Journal of Orthopaedic Research 1: 22-29.
Woo S, Gomez M and Sites T (1987): The biomechanical and morphological changes in the medial collateral ligament of the rabbit after immobilization and remobilization. The Journal of Bone and Joint Surgery 69A: 1200-1211.
The therapist should always determine the phase of the inflammatory process before starting treatment. Acute traumatic injuries often present with a clear time of onset, but with overuse injuries, because of the insidious onset, it is more difficult to identify the current phase. Continuous reinjury as is common with overuse will repeatedly set back the progression of the inflammation phase, and therefore it is impossible to tell when the inflammatory process started. Because of this the therapist should always, unless proven otherwise assume that it is in the vascular inflammatory phase (Hujibregts and Smith 1999). Treatment of acute and chronic ligament/tendon injuries can be confusing because of the fine balance between adequate rest and preventing atrophy of surrounding muscles. This has led to the development of very detailed exercise programs of for example ACL, Achilles- and Patella tendon (Hyman and Rodeo 2000).
Healing of connective tissue
During the inflammation phase the tensile strength is dependant upon fibrin, which is very fragile, and tension could easily disrupt it. In order to control it is essential to limit the amount of post-injury oedema using ice, elevation compression. Other forms of management are NSAID´s and modified rest. Ultrasound and laser has shown to accelerate the inflammation process (Hunter 1994, Reid 1992).
In the regeneration phase it is important to maximize strength, the collagen fibres should relate to the undamaged tissue. Careful tensioning during this phase promotes correct alignment by remodelling the collagen fibres. The rate of increase in wound strength is proportional to the amount of collagen laid down (Hunter 1994).
Management in the remodelling phase is to minimize immobilization, balance functional stresses with increasing tissue strength, re-establish ROM and enhance proprioception (Reid 1992).
Prevention of scar tissue and contraction and promoting the correct orientation of collagen fibres is essential for the best functional outcome. The wound should be subjected to tension on a regular basis to promote extensibility of the scar tissue (Hunter 1994).
Modalities and pharmacology
There has been a great deal of research on the ability to influence the basic biology of tendon healing using pharmacological and physical modalities. Low intensity laser beam (photo stimulation) and electric stimulation has been shown to enhance tissue healing the research showed an increased total collagen production, but there were fewer cross-links per collagen molecules, the mechanism remains unclear. It is suggested that it promotes nucleic acid synthesis and cell division of human fibroblasts. Ultrasound has been reported to improve scar maturation and to decrease inflammatory cell infiltration into healing flexor tendon (Hyman and Rodeo 2000). A combination of ultrasound, electrical stimulation and laser stimulation was found to increase collagen synthesis and tensile strength in healing rabbit’s tendons (Gum and Reddy 1997).
Cold is useful in the immediate care of an acute tendon injury to decrease inflammation, oedema and pain. Heat has been shown to increase chemical activity, metabolic rate and tissue extensibility and decrease muscle spasm, pain and ischemia. Heat should not be used in the acute phase of the inflammation (Józsa and Kannus 1997) .
Various growth factors have been found to effect cell proliferation, matrix synthesis, and cell migration in intrasynnovial and extrasynnovial tendons, the most promising are (TGF, IGF-1 and PDGF). Ligaments treated with PDGF, IGF-1 and FGF had increases in strength, stiffness and breaking energy (Hyman and Rodeo 2000).
The current opinion on the use of NSAID´s in acute tendon strains and partial tendon ruptures should be administrated during the inflammation and proliferation phase (Józsa and Kannus 1997).
Tensile loading/stress
In the early stages of rehabilitation it is of great importance that the injured tissue is well protected. Maintaining the blood supply is critical to prevent necrosis of the collagen, and this may be a problem in areas with compromised vascularity because of friction, compression or torsion, for example achilles and supraspinatus tendon. It is also important to have a regime of progressive tensile stress, which will help the fibrillar alignment, appropriate cross-linking and normal mechanical properties, during subsequent phases. Research shows that exercise has a positive effect on mechanical and morphological characteristics of the injured tissue. There is a problem with applying a progressive regimen of tensile force to strengthen the tissue without reinjuring it (Hujibregts and Smith 1999).
Deep friction can in early phases enhance the mobilisation of tissue fluid, and in later stages stimulate fibre orientation, prevent cross linking and adhesions and increase blood flow.
Stretching should be used with increased tissue temperature and slow prolonged stretching to gain a permanent increase in ROM (Reid 1992).
Ligaments
The rehabilitation of ligaments depends on if the joint is stable or not, unstable joints may need bracing techniques or surgery. Injured ligaments have abnormal proprioception, which makes the proprioceptive training another factor to include in the rehabilitations program.
Joint position sense (JPS) in ACL deficient knees has been described as impaired, although knee stability can improve with exercise therapy, there may be no improvement in JPS. The role of JPS in the stability of ACL deficient knees remains unclear (Hyman and Rodeo 2000).
Heavy load eccentric calf muscle training for chronic achilles tendinosis
A study on eccentric training was conducted with 15 middle-aged recreational athletes, who had the diagnosis chronic achilles tendinosis (degenerative changes) and where selected for surgical treatment. They also had a control group with the same diagnostic criteria that underwent surgical treatment. The training group followed an eccentric training program 2 times a day for 12 weeks. The program included eccentric calf muscle training with both the knee extended and flexed (3×15 for each exercise). The patients were told to go ahead with the exercise even if they experienced pain. When they could perform the exercise without pain they were instructed to increase the load.
After 12 weeks the training group were all back to penury level. The control group were all back to pre-injury level after 6 months.
Possible explanations may be either the effect of stretching with a lengthening of the muscle-tendon unit, and consequently less strain during ankle joint motion. Or loading within muscle tendon unit with hypertrophy and increased tensile strength in the tendon, thus remodelling is induced from eccentric loading (Alfredson and Lorentzon 2000, Alfredson et al 1998).
Ligament or tendon injury occurs when the load acting on the tissue is over its own capacity. When the load is over phase III in the above-mentioned stress-strain curve, tissue failure occurs. As soon as the ligament starts to heal up after the injury, usual healing process will proceed namely: 1. inflammatory; 2. cellular and matrix proliferation (regeneration); and 3. remodeling.(Hildebrand and Frank 1998, Reid 1992)
In the following, changes in the tissues structures as well as their mechanical properties during the healing processes will be discussed.
A. Healing response within tendon and ligament toward injury
I. Inflammatory phase
At the cellular level, alteration of structural makeup starts.
• Formation of fibrin clot (Hildebrand and Frank 1998, Reid 1992).
• Fibroblasts proliferation (Hayman and Rodeo 2000, Liu et al 1995, Reid 1992).
• Ligament: High Type III collagen (loosely packed thin fibrils), low Type I collagen (densely packed thick fibrils)
Tendon: Similar amount of Type I & III production (Liu et al 1995).
II. Regeneration
At cellular level:
• Type I collagen fibrins bridge up the gap between the torn ends (Hildebrand and Frank 1998).
• Spare collagen framework formed (Reid 1992).
• Formation of scar with Type I collagen comprises large percentage of scar tissue (Frank 1996, Liu et al 1995).
• Fibroblast remains predominant cell.
III. Remodeling
1. At cellular level:
• There is an increase of collagen density & cross link, alignment of collagen fibers in the axis of ligament (Hayman and Rodeo 2000, Liu et al 1995).
• Histological normal ligament after 7 months, mechanical properties still not fully returned to normal state.
• Still high type III collagen percentage in repaired ligament (Liu et al 1995)
• Alignment of collagen fibers is longitudinally along lines of stress in tendon, minimal histological difference from normal tendon after 20th week.
2. Mechanical properties of an healed injured tendon and ligament
• The tensile strength shows dramatic reduction (Reid 1992).
• Ligamentous strength after repair is in the region of 60 to 70% normal after 6 weeks of healing (Reid 1992).
• It takes up 3 months before 80% of the original strength is acquired (Reid 1992).
• Intra-articular ligaments usually gain tensile strength more slowly, 3 months 50% of normal strength & 6 months 70% functional strength. (Reid 1992).
• Strength and stiffness restore to 40 – 90% of normal values in animal studies, only about 30 – 70% of the material strength has returned (Frank 1996).
• Viscoelastic properties appear to return to 70 – 90% of normal values (Frank 1996).
• Ligament (using MCL) heals structurally 70 – 80% of the strength and structural stiffness of a normal MCL. The scar reaches a maximum of only about 30% of normal MCL strength, even after months to years of healing (Frank et al 1999, Frank 1996).
3. Formation of scar tissue
• Scar development: inflammation and formation of granulation tissue, scar proliferation and scar remodeling (Frank 1996).
• Comparison between ligament scar and normal ligament properties (Table 1.)
Normal ligament
Ligament Scar
Collagen aligned
Collagen disorganized
Collagen densely packed
Defects between collagen fibers
Large collagen fibrils
Small collagen fibrils
Mature fiber cross-links
Immature cross-links
Primarily collagen type I (< 10% type III)
More collagen type III
Small proteoglycans
Some large proteoglycans
Other components minor
Excesses of other components
Cell metabolism low
Cell metabolism high
Low cell density
Increased cell density
Low vascularity
Increased vascularity
• 3 major reasons for scar weakness – “flaws”, smaller than normal collagen fibril sizes and abnormal collagen cross-linking (Frank et al 1999, Hayman and Rodeo 2000).
• Normal ligaments Densely packed nearly parallel arrangements of a range of sizes of collagen fibers, cross-links are stable and resistant to breakdown. However the scar tissue has poorer cross-links and needs months to realign and abnormal collagen cross-linking (Frank et al 1999, Hayman and Rodeo 2000).
B. Effects of Immobilization and Mobilization
• Immobilization protects some ligament repairs grossly, causes isolated ligament scars to be less stiff and significantly less strong than scars in the joint that have been allowed to move, decreases ligament strength, has potential minimization of scar length which leads to ligament laxity (Frank 1996).
• Study in rabbit’s ACL showed changes in the shape and intracellular structure of fibroblasts from the ligament after immobilization, ligament switches progressively from an anabolic to a more catabolic state (Boorman et al 1998).
• Mobilization with some degree of mechanical load appears to be essential to the normal maturation and maintenance of the structural and mechanical properties of ligaments (Boorman et al 1998).
• Mobilization with controlled movement has been shown to improve scar stiffness and strength without compromising scar length, shown to stimulate collagen synthesis, matrix remodeling, production of better quality scar when compared with those produced with immobilization, production of increased scar mass to resist the tensile stresses involved (Frank 1996).
• Movement creates tension that increases fibroblast proliferation, migration and collagen synthesis, aligns the fibroblast & collagen fibrils parallel to the direction of the force. Therefore a healing ligament will have a high tensile strength (Liu et al 1995).
• In tendon, passive mobilization can prevent adhesion between the sheath and the healing tendon that restricts motion.
• In conclusion, mechanical stimulus (mobilization) has a significant affect on ligament and tendon structure.
Overview on management of tendon / ligament injuries
Grade I injury
Signs
Management
– Minimal loss of structural integrity
– Minimal functional loss
– No abnormal motion
– Early return to training – some protection may be necessary
– Little or no swelling
– Localized tenderness
– Minimal bruising
Grade II injury
Signs
Management
– Significant structural weakening
– Tendency to recurrence
– Some abnormal motion
– Need protection from risk of further injury
– Solid end feel to stress
– May need modified immobilization
– More bruising and swelling
– May stretch out further with time
– Often associated hemarthrosis & effusion
– Dramatic reduction of tensile strength, a ware of the dangers of unduly stretching healing structures at this point.
First category
– minimum tearing of a few fibres
– some pain
– little loss in structural integrity, no joint instability felt clinically
Second category
– moderate tearing of collagen fibres
– severe pain
– some loss of structural integrity, joint instability can be detected clinically
Third category
– most of the collagen fibres have ruptured, near complete tear
– severe pain
– joint is completely unstable
–
Factors affecting the Mechanical Properties of Ligament
Factor: Maturation and Aging
During maturation (up to 20 years of age),
-the number and quality of cross-links with the collagen molecules increases
-collagen fibril diameter increases
There is an overall increased tensile strength and stiffness of the ligament. After maturation, with aging, there is a gradual decrease in strength and stiffness.
In one study, the ACL was found to exhibit age-related changes. The young (16-26 years) adults’ are considered to be 3 times stronger than those of older adults (48-86 years), 1.7 times for the elastic modulus i.e. stiffer and 1.5 times for failure strain i.e. ultimate load (Panjabi and White, 2001).
Factor: Activity, Exercise and Disuse
Activity/ Exercise
Ligaments appear to remodel and become stronger and stiffer when there is mechanical demand on them (Noyes et al, 1974). Physical training improves the tensile strength and also the ligament-bone interface (Woo et al, 1983). Daily activity is sufficient to maintain 80-90% of a ligament’s mechanical potential. It has been found that injured ligaments that are exercised have significantly better physical and biomechanical properties than those not exercise (Nigg and Herzog, 1999).
Disuse/Immobilisation
In contrast, immobilization and disuse, decreases the tensile strength and stiffness of ligaments. In a study by Woo et al(1987) on rabbit knee, after 9 weeks of immobilization there was a corresponding 69% decrease in ultimate load and an 82 % decrease in energy to failure. Biomechanically, the water content decreases, collagen mass is reduced whilst the number of cross-links is increased and the ligament becomes stiffer.
Immobilization has a more immediate and substantial effect compared to effects of exercise. Upon remobilization, the effects of immobilization are reversible but considerable time will be needed to regain the former strength and stiffness. Noyes (1977) experimented 8-weeks immobilization on animal ligaments and found that 12 months of re-conditioning was necessary before it was comparable to initial values.
Factor: Pregnancy and Postpartum Period
Rundgren (1974) reported that the decreased stiffness of tendons and ligaments during early postpartum was restored later.
Factor: Temperature
Generally, ligaments become stiffer with cold and have increased extensibility with heat.
Factor: Ligament differentiation- Properties
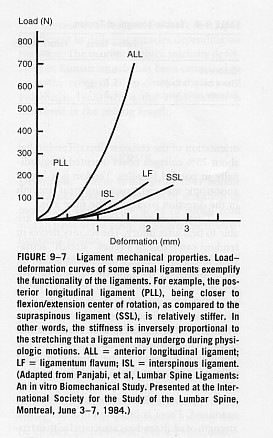
-Though ligaments follow a general stress-strain curve, the diagram below demonstrates that different ligaments exhibit different degree of stiffness and elasticity, hence different failure properties to injury. Posterior Longitudinal ligament is the stiffest and the most flexible ligaments are interspinous and supraspinous ligaments (Panjabi and White, 2001).
-Differences exist between ACL and MCL in their biologic responses during its midsubstance injury. MCL heals quickly and predictably even without repair whilst ACL fails to establish a significant healing response even if direct repair is done. There are ultrastructural differences. The cells of MCL are similar to those of fibroblasts (synthesis of collagen) but the ACL resemble fibrocartilage cells.
Factor: Diabetes Mellitus
Diabetes Mellitus is a disturbance of the normal insulin mechanism. When compared to nondiabetics, diabetics show a higher rate of tendon contracture (29% vs 9%), tenosynovitis (59% vs 7%), joint stiffness (40% vs 9%) and capsulities (16 % vs 1 %).
• Osteotendinous junction (trauma/overuse or disease)
• Mid tendon (often occurs in degenerative tendons – tendinosis)
• Musculotendinous junction (vulnerable point, common site of tendinitis)
Tensile strength of a healthy tendon is normally more than twice the strength of its muscle and stronger than compact bone. A tendon with a cross sectional area of 1cm² is capable of supporting a weight of 500 – 1000kg (Jozsa and Kannus 1997). Therefore unless the tendon is unhealthy it will normally breakdown at the MTJ. The MTJ has a delicate ultrastructure and is often the weakest point of the muscle-tendon unit making it more susceptible to injury (Jozsa and Kannus 1997).
Tendon Injury Patterns
• Acute
• Chronic
Acute tendon strains
Excessive loads greater than 4 -10% of original length or direct contusion will cause acute injury to tendons. Under normal physiological circumstances tendons function in the toe and linear region of the stress-strain curve. Loads greater than this will cause damage most often at the MTJ. Muscles that cross two joints are predisposed – hamstring, rectus femoris, gastroc. This normally occurs when lengthening of the muscle combines with eccentric contraction.
Chronic/overuse tendon injury
Repeated loading has been shown to cause overuse tendon injuries when the destructive forces of applied stress exceed the reparative process. This occurs when the tendon is strained repeatedly to 4 – 8% of its original length until it is unable to endure further tension, where upon injury occurs (Hyman and Rodeo 2000).
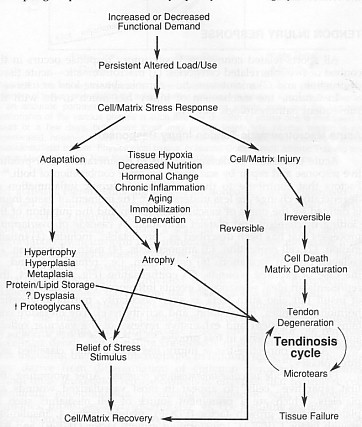
This causes micro and macroscopic injury to collagen fibrils, noncollagenous matrix, and microvasculature causing inflammation. It is currently believed that the earliest pathophysiological changes occur in the paratendon (surrounding the tendon) causing paratendinitis. Continued microtrauma whilst in the inflammatory stage will lead to proliferation of synovial cells, fibroblasts and capillaries leading to fibrosis and thickening of the paratendon. Thus, tendonitis and paratendinitis are the earliest clinical manifestations of overuse tendon injury. If the damage progresses, tendinosis, partial tears and complete rupture may ensue (Jozsa and Kannus 1997).
Classification of Injury
Paratendinosis is inflammation of the paratendon only, either lined by synovium or not. It can be acute or chronic. Signs and symptoms: swelling, pain crepitus, local tenderness, warmth, and dysfunction.
Tendinitis is symptomatic degeneration of the tendon with vascular disruption and inflammatory repair response. Symptoms are inflammatory and proportional to the vascular disruption. It can be acute, subacute or chronic.
Tendinosis is when the tendon undergoes intratendinous degeneration due to cell atrophy, affecting its mechanical properties. The result of the pathological changes is decreased tensile strength that predisposes the tendon to injury. It is often asymptomatic since it is non inflammatory. It can be defined as “a change in tissue from a higher to a lower or less functionally active form”(Leadbetter 1992).
(Leadbetter 1992) illustrating Achilles paratendonitis +/- tendinosis
Classification of Tendon injuries and disorders
intratendinous degeneration due to atrophy (aging, microtrauma, vascular) can be nonsymptomaticsymptomatic degeneration of tendon with vascular disruption and inflammatory repair response
NEW
OLD
DEFINITION
Paratendinitis
Tenosynovitis
Tenovaginitis
Peritendinitis
inflammation of only the paratendon either lined by synovium or not.
Paratendinitis with tendinosis
Tendinitis
paratenon inflammation associated with intratendinosis degeneration
Tendinosis
Tendinitis
intratendinous degeneration due to atrophy (aging, microtrauma, vascular) can be nonsymptomatic
Tendinitis:
Tendon strain/tear acute, subacute chronic
symptomatic degeneration of tendon with vascular disruption and inflammatory repair response
Adapted from (Leadbetter 1992)
Factors contributing to tendon injury (extrinsic/intrinsic)
• Excessive or Repeated loads – causes:
Poor biomechanics, joint laxity, leg length discrepancy – will alter direction, speed and size of force. e.g. foot hyperpronation has been linked to Achilles tendon injuries.
Muscle imbalances – weak muscle, tight muscle, poor timing of muscles can alter force distribution leading to strains.
Muscle fatigue, poor flexibility, – may increase load on tendons
Overweight – increased load
Training errors, poor equipment – e.g. prolonged exercise duration, could increase load
• Vascularity – if the tendon has a reduced blood supply (hypovascularity) it can lead to tissue hypoxia resulting in tendon cell atrophy contributing to tendon degeneration that results in tendinosis.
Hypovascularity has been linked most frequently to explain hypoxic degenerative tendon changes (O’Brien 1992). (Archambault et al 1995) reported that blood supply is irregular to the TA tendon mid-portion and at sites of twisting or around bony prominences. This study reported decreased vascularity in the TA tendon 4cm proximal to its calcaneal insertion. He suggested the cause was due to intermittent ischaemia during exercise – with subsequent reperfusion – leading to influx of oxygen derived free radical – these are highly reactive molecules that have a role in phagocytosis leading to tissue self destruction/degeneration.
Secondly, hypovascularity is linked to calcifying tendinopathy – persistent hypoxia may transform tenoblasts into chondroblasts that function anaerobically. Results in calcium deposition within the tendon.
Thirdly, regions of the tendon with poor vascularity will be less likely to cool over heated tissue caused by muscle activity, leading to exercise induced hyperthermia. This study reported that the intratendinous temperature of superficial digital flexor tendons of exercising horses (43 – 45º) exceeds the temperature at which fibroblast cell death occurs (42.5º). Thus, avascular regions of tendons may lack the ability to cool down the tissue leading to hyperthermic damage during exercise (Wilson and Goodship 1994).
• Genetic disorders/disease/infection – genetic collagen disorders like Ehlers-Danlos syndrome results in less cross-linking leading to weaker tendons. Rheumatoid diseases causes gross structural changes to tendons. Penetrating wounds can lead to tendon infections.
• Aging – results in stiffer, less compliant, weaker tendons with a reduced repair ability increasing their susceptibility to injury. This is due to decreased elastin, water and proteoglycan content, reduced collagen resynthesis and reduced vascularity and tenoblast activity (Hyman and Rodeo 2000).
• Endocrine factors – Diabetes Mellitus results in a compromise to tendinous microcirculation which may lead to tissue anoxia, cell necrosis thus predisposing a diabetic tendon to chronic paratendinitis or tendinosis (Jozsa and Kannus 1997). Hormonal changes in women can reduce the levels of estrogen, which has been linked to decreased collagen production.
• Nutritional deficiencies of cofactors (vitamin A, C and copper) important in collagen synthesis and cross-linking may affect tensile strength of tendons.
• Inactivity/Immobilization – (Jozsa and Kannus 1997) studied the effect of immobility on a human MTJ. Results showed the contact area between muscle cells and tendon collagen fibrils decreased to almost 50% in 3 weeks. There was also an increase in the amount of type III collagen at the MTJ, which is weaker than type 1 and a decrease in the amount of tenascin, an adhesive protein found in the MTJ. The overall effect will be to decrease tensile strength.
• Medication – corticosteroids (oral/injections) and certain antibiotics (flouroquinolone) can lead to pathology of the cell matrix. Excessive use of corticosteroid injections has been linked to tendon rupture (Jozsa and Kannus 1997). This may be due to its anti-inflammatory effect, as the injection reduces the pain, the patient will resume normal activity on the weak, degenerative tendon which could result in rupture.
• Skeletal maturity – osteotendonopathies like Osgood-Schlatters disease at the tibial tuberosity or Severe’s disease at the calcaneus can occur in the immature child due to tensile overload at the apophysis. Avulsions of small areas of the ossification center are found.
The flow diagram illustrates how tendinosis results from a failed cell matrix adaptation to excessive changes in load use. (Leadbetter 1992)
There are three major types of behaviour characteristic of viscoelasticity. Firstly creep, this is increasing deformation under constant load in the elastic portion of the stress-strain curve (Sandrey 2000). So that with a constant load, collagen length will increase over time. (Huijbregts and Smith 1999).
There are 3 major regions of the stress-strain curve that are of significance:
Mechanical Properties of Ligaments and Tendons (Jozsa and Kannus 1997)
I. Toe region:
As the load increases so does the recruitment of collagen fibres causing them to ‘uncrimp’. This occurs when collagen is stretched to approximately 2% of its original length and returns to normal length when the force is removed, thus it is within its physiological range. It is characterized by relatively low stiffness. There is a non-linear relationship on the stress-strain curve at this stage.
II. Linear region (Elastic phase):
As the collagen fibrils become gradually uncrimped, the fibril itself is being stretched. There now becomes a linear relationship between deformation and load, as the tissue becomes relatively stiffer. This occurs when collagen is stretched to 2-4% of its original length and returns to its original geometric shape. The tissue is said to be elastic.
III. Yield and Failure region (Plastic phase):
The continued increase in load past 4% causes microfailure to the fibrils and damage to cross-links. It results in a plateau effect on the curve: this point represents the ultimate tensile strength of the tendon and is termed the ‘yield point’. The yielding of fibres occurs when the deformation is approximately 4-10% of the resting length. Stiffness is reduced and the fibrils do not return to normal length on release, the tissue then becomes ‘viscous’. This is known as ‘plastic’ deformation.
Finally, complete failure occurs as the ligament/tendon ruptures. Obviously this is a non-physiological range and there would be an inability to support load or function (Huijbregts and Smith 1999, Jozsa and Kannus 1997, Reid 1992).
Collagen demonstrates various mechanical and physical properties in response to load and deformation to allow it to withstand high tensile stresses. The point between the elastic and plastic region is where gross integrity is disrupted.
Viscoelasticity
Tendons and ligament are viscoelastic materials and display sensitivity to different strain rates. Viscoelasticity indicates collagen’s property of demonstrating time dependent and variable elastic behaviour. Thus, the relationship between stress and strain is not constant but depends on the time of displacement or load.
There are three major types of behaviour characteristic of viscoelasticity. Firstly creep, this is increasing deformation under constant load in the elastic portion of the stress-strain curve (Sandrey 2000). So that with a constant load, collagen length will increase over time. (Huijbregts and Smith 1999).
The second behaviour characteristic is force relaxation. This can be defined as the decrease in the amount of force required to maintain a set amount of deformation over time (linear region of curve). This occurs because the tissue relaxes (stress relaxation).
The third characteristic is hysteresis or energy dissipation. This means that if a viscoelastic material is loaded and unloaded, the loading curve will not follow the loading curve. The difference between the two curves represents the amount of energy that is dissipated or lost during loading. It refers to the amount of relaxation the tissue demonstrates during single cycle of deformation and relaxation. It is therefore an indication of the viscous properties of the tissue.
Description
Both ligaments and tendons are fibrous, dense regular collagenous connective tissues. Ligaments join bone to bone in a joint whilst tendons join muscles to bone.
Function of Ligament
1. to resist external load. Prevents excessive motion
2. to guide joint motion. Therefore, guide relative movements of the 2 bones
3. to passively control the maximum range of movement. Therefore, augment mechanical stability of the joint. Hence, the ligament length, stiffness, location and orientation with respect to the articulating bones affect the motion patterns.
4. to provide motor control
Function of Tendon
1. transmit tensile forces from muscle to bone
2. advantage mechanical pulley enables muscle belly to be an optimal distance from joint without requiring an extended length of muscle between origin and insertion.
3. to maintain body posture
4. to provide motor control
Hierarchical Structure
In the diagram (Reid, 1992) below depicts the structure of either the tendon or ligament . They are split into smaller fascicles and each fascicle contains the basic fibril and the fibroblasts. It is the crimp/waviness of the fibril that contributes significantly to the non-linear stress-strain relationship of ligaments and tendons. The fibroblasts are the biological cells that produce the ligament or tendon.
Contents
Collagen
Both tendons and ligaments are made up of mainly 70% water, 20% collagen and about 2% of ground substance. However, collagen contributes to 70% of its dry weight. Collagen is a fibrous protein with high tensile strength.
However, the arrangement of collagen fibres differs between tendons and ligaments. Ligaments are organized in a dense but random/loose-packed network unlike tendon that have an orderly, parallel, linearly packed arrangement. This facilitates tendons to handle high unidirectional (uniaxial) tensile loads during activity whilst ligaments generally sustain tensile loads in a predominant direction but also bear smaller tensile loads in other directions.
A. Ligaments- densely packed collagen in a random-packed network but towards a general primary direction.
B. Tendons- linear arrangement(Reid, 1992)
Tendons and extremity ligaments have very little protein elastin. Ligament have a higher ratio of fibrillar Type I collagen to Type III collagen (elastic). Thus, ligament’s mechanical properties provide it with not only the flexibility but also the tensile properties to resist force.
Ground substance
It provides mechanical properties in compression. Ligaments buckle under compression and do not provide any support.
Innervation and mechanoreceptors
Within tendons there are 4 types of nerve endings: type I or Ruffuni corpuscles monitor pressure; two types of mechanoreceptors – type II or Vater-Pacini corpuscles and type III or Golgi tendon organs; type IV free nerve endings monitor pain. Together they provide essential afferent sensory neural information that controls the movements of the body via the central nervous system (Jozsa and Kannus, 1997). The ACL is richly innervated by mechanoreceptors (Schultz et al, 1984). It has been proposed that the sensory information provides neuromuscular control and hence, contribute to dynamic stability of the joint.
Outer structure and Insertion into Bone
Both tendons and ligaments are surrounded by loose areolar connective tissue. Whilst this tissue around ligament is not significant, in tendons, this tissue known as paratenon is more structured and forms a sheath that protects it and promotes gliding.
Clinical implication: introduction of passive motion for healing/repair to prevent adhesion between the sheath. (study of 3 groups of dogs-complete immobilization; delayed immobilization and early passive motion. The latter group’s tendon strength and motion improved more quickly than the rest.)
Load refers to the external force applied to the collagen. Stress refers to the amount of resistance within the collagen to the load. Strain is the amount of deformation that occurs in the collagen because of the load (Sandrey 2000) .
According to Gray’s Anatomy (Williams 1995, page 88-90), the general connective tissues can be classified, in relation to their cell types and their extracellular matrix, into irregular and regular connective tissue.
The irregular tissue can be further divided into:
• Loose tissue. Loose connective tissue is ubiquitous. It forms the subcutaneous tissue of eyelids and external reproductive organs, the submucous coat in the digestive tract and the subserous tissue. It is found between muscles, vessels, nerves and interior organs. Interweaving in all directions, thin collagen and elastin fibres form the basis of this tissue. The casual-straggled arrangement of the fibres into the loose connective tissue determines elasticity and tensile stamina.
• Dense tissue. This tissue constitutes the sheaths of organs, which need protection from mechanical stresses. The extracellular matrix encloses thick bundles of collagen fibres setting in three dimensions and providing substantial strength. It is found in structures such as the reticular layer of the dermis, the sheaths of muscle and nerves and the adventitia of large blood vessels, the capsules of different glands, the coverings of diverse organs, the sclera of the eye, the periostea and the perichondria.
• Adipose tissue. This tissue looks like loose connective tissue, but it contains many more fat cells. It gathers in certain regions such as subcutaneous tissue, and around the kidneys, in the mesenteries and omenta, in the female breast, in the orbit behind the eyeball, in the marrow of bones, deep to the plantar skin of the foot, and as localized pads in many joints.
The regular connective tissue is predominantly consists of collagen fibres with some elastin fibres, which are located between them, except some ligament such as ligamenta flava, which are rich in elastic fibres. Regular connective tissue comprises tissue with regularly oriented fibres such as fasciae, aponeuroses, ligaments and tendons. The fibres in those structures are oriented towards stress forces. They keep their helical form, which enhances their mechanical features.
According to Gray’s Anatomy (Williams 1995, page 75), structural connective tissues are conveniently divided into:
• General or ordinary types, which is widely distributed,
• Special skeletal types, namely cartilage and bone, and
• Haemolymphoid tissue and their precursors
The following paragraphs will describe the general connective tissue, because the description of the other types of connective tissue is out of the purpose of this text.
General connective tissue
Connective tissue is formed of cells and extracellular matrix (fig.1). The matrix in turn is composed of fibres and ground substance. The ground substance is a jelly, aqueous environment. This environment contains collagen fibrils and is controlled by specifically attached proteoglycan molecules. Haemal tissue of the bone marrow or lymphoid tissue provides connective tissue with a number of cell types. The cells reach the connective tissues by the circulating blood and migrate into them through their endothelial walls (Williams 1995).
fig. 1. Areolar connective tissue (Taken from www.google.com)
According to Gray’s Anatomy (Williams 1995, page 75-80), cells of general connective tissue can be separated into:
1. The resident cell population (fibroblasts, adipocytes, persistent mesenchymal stem cells, etc)
2. The immigrant population. They are wandering cells with various defensive functions (macrophages, lymphocytes, mast cells, neurotrophils and eosinophils).
1. The resident cell population.
Fibroblasts are usually the most numerous cells. They synthesize and secrete most of proteins of the extracellular matrix of connective tissue. They transform into fibrocytes when they become old and inactive.
Fibroblasts lay down collagen and elastin fibres and adhere to them. During wound repair, they are predominantly active by multiplying and laying down a fibrous matrix. Various factors such as steroid hormone levels, dietary content and mechanical stress affect their activity. Vitamin C deficiency leads to an impairment of collagen formation.
Adipocytes (lipocytes, fat cells) exist singly or in groups in the meshes of many but not all connective tissues. They are specially numerous in adipose tissue.
Pigment cells occur in the dermis of skin and in the iris and choroid of eye. Their function is generally to prevent light from reaching adjacent cells.
2. The immigrant cell population
Macrophages (histocytes) are also numerous in connective tissues, where they may be either attached to matrix fibres (stationary or fixed macrophages) or they may be motile (nomadic macrophages). These cells are important phagocytes, which are particularly active during inflammation.
Lymphocytes. These cells are typically present in small numbers, but are numerous in pathological states, migrating from adjacent lymphoid tissue or from circulation.
Mast cells (histaminocytes) are important defensive cells, which occur particularly in loose connective tissues and often in the fibrous capsules of some organs such as liver. They are associated with the inflammation. They migrate to the injured tissue where they may be disrupted to release their contents such as histamin, heparine. The signal for this migration is the direct mechanical or chemical trauma or the contact with specific antigens to which the body has previously been exposed.
Neutrophil and Eosinophil leucocytes. They frequently present in small numbers but when the tissues are infected, they migrate to them in increased numbers.
Extracellular matrix
The presentation of the ECM will concentrate on those elements that have prominent mechanical functions. In this sense, the ECM consists of 3 types of macromolecules – fibres (collagen and elastin), proteoglycans (PGs) and glycoproteins. Each of these macromolecules is synthesized and maintained by cells, which are specific to the tissue type (Culav et al 1999).
1.Fibres.
The two most important fibrous components of the ECM are a) collagen and b) elastin, which are insoluble macromolecular proteins. The most prominent feature of collagen is their ability to resist tensile loads. Generally, under tension, they show minimal elongation (less than 10%). This is not the true elongation of the individual fibres. It is the result of the straightening of fibres that are crowded in various 3-dimensional arrangements. Conversely, when a force stretches elastic fibres, they may increase their length by 150%, and after the force has been removed, they return to their previous configuration (Culav et al 1999).
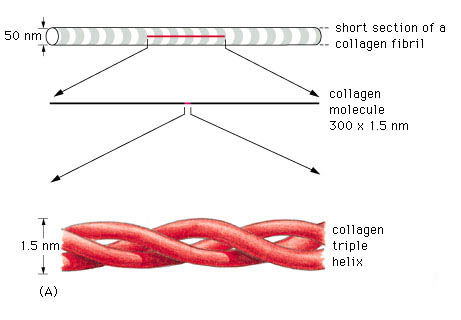
a) Collagen. Collagen is the most abundant protein in the human body, accounting for nearly 30% of all protein (Williams 1995). Nineteen different types of collagen fibres have been found by 1999, all with individual characteristics that serve specific functions in a variety of tissues (Liu et al 1995, Mayne 1997, Culav et al 1999).
The “collagen family” is divided into four subclasses (Burgeson et al 1992, Williams et al 1997):
• Fibrillar collagens (type I, II, III, V, XI)
• Basement membrane-associated collagen (type IV, VII)
• Fibre-Associated (type IX, XII)
• Short-Chain (type VIII, X)
The following paragraphs will describe the fibrillar collagen types I, II and III, which are among the most abundant proteins in the body and are the most relevant to physical therapists.
The fibrillar collagens are estimated to constitute over 70% of the total collagen found in the body (Culav et al 1999). Collagen type I has the broadest distribution (Liu et al 1995, Hukins 1990).
Fig. 2 Collagen triple helix (Taken from www.google.com)
A triple helix is the general structural feature that identifies the fibrillar collagens (fig.2). The diameter and length of the fibrillar collagens vary. Their surfaces have characteristic striations formed by gaps between the ends of the molecules (fig.3) (Williams 1995).
fig.3. Striations of collagen fibres (Taken from www.google.com)
Type I collagen account for 60% to 90% of the dry weight of skin, ligaments, tendon and bone (demineralised) (Mayne 1997, Williams et al 1997). It is also found in many thin tissues including the lungs and dentin and the sclera of the eyes. In addition, it is the major constituent of mature scars (table 1) (Williams et al 1997). Furthermore, it is found in annulus fibrosus of the intervertebral disc, fibrous cartilage and fascia (Liu et al 1995, Culav et al 1999).
Type II collagen accounts for more than half the dry weight of hyaline and elastic cartilage, and it is found in the nucleus polposus and in the vitreous gel of eye (table 1). It is also transiently present in many tissues during embryonic development (Williams et al 1997, Liu et al 1995, Culav et al 1999).
Collagen type
Tissue distribution
Ultra-structure
Site of synthesis
Function
Type I
Dermis, bone, tendon, teeth, fascia, sclera, organ capsules, fibrous cartilage
Densely packed thick fibrils with marked variation in diameter
Fibroblast, osteoblast, odondoblast
Resistance to tension
Type II
Hyaline and elastic cartilage, intervertebral disc, vitreous
No fibres; very thin fibrils embedded in abundant ground substance
Structural maintenance in expansile organs; wound healing, mediate attachments of tendon, ligament and periosteum to bone cortex
Table 1. Adapted from Liu et al (1995)
Type III collagen is abundant in large blood vessels, skin, liver, spleen, and is found in small amounts in most tissues that contain type I collagen (table 1), but it is not present in bone (Williams et al 1997, Liu et al 1995). For example, the dermal collagen is constituted of 80% – 85% of type I collagen and 15% – 20% of type III collagen (Williams et al 1997). It is the earliest collagen, which is laid down in the healing process, and because its cross-links can be generated rapidly, they provide mechanical strength to the newly synthesized matrix. As the healing tissue gains in strength, the stronger type I fibres replaces type III fibres, if the injured tissue is tendon or ligament (Culav et al 1999). But, if the injured tissue is cartilage, type II fibres replace them.
Biosynthesis of fibrillar collagen
The initial transcript of the major fibrillar collagens (types I, II, and III) is secreted from the nucleus of the cell as interstitial collagen (fig.4). The biosynthesis of individual chains then occurs on the rough endoplasmic reticulum. Subsequently, the chains are folding in the individual triple chains to form the collagen molecule, which is named procollagen. Then, the collagen molecules move to the Golgi apparatus, by which they promoted and extruded from the fibroblast (Mayne 1997, Burgeson et al 1992).
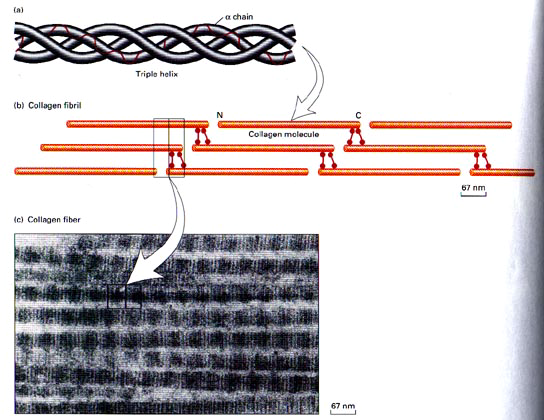
Before fibril formation can occur, the procollagen molecules lose their noncollagenus extensions at their terminal regions and become negatively charged molecules named tropocollagen. Many molecules of the tropocollagen are arranged side by side to form the quarter-staggered fibrils. This configuration is visible in the electron microscope and characterized by distinct cross-band. The fibrils are stabilized by the formation of cross-links between individual molecules (Mayne 1997).
The fibrils join together to form fibres (fig.4), which in turn combine together to form bundles of collagen fibres The first can be seen only under the light microscope, while the bundles can be seen unaided (Culav et al 1999). This procedure occurs for the morphogenesis and remodelling of tendons. For the other tissue, it is poorly understood how collagen fibrils form and whether fibril formation is closely associated with the cell ( Mayne 1997). Fibrils may also be formed of more than one type of collagen (Culav et al 1999). For example “The banded cartilage collagen fibres are thought to contain types II and XI, and the noncartilage banded fibres contain mixture of types I, III, and V collagens.” (Burgeson et al 1992).
In tendons, the majority of fibres have parallel alignment. In ligaments, the collagen fibres are positioned in slightly less parallel arrangement. In skin, fibres are arranged in random orientation. Collagen plays an important role in attaching tendons and ligaments to bone. (Culav et al 1999)
fig.4. Biosynthesis of fibrillar collagen (Adapted from Culav et al 1999)
b) Elastin.
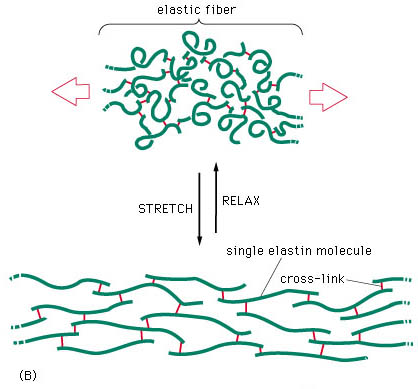
The presence of rubber-like elastic fibres determines the elastic properties of tissues such as skin, large blood vessels, lung and large ligaments (fig.5). Elastic fibres are unstructured, in the sense that their molecular components are not assembled in the regular pattern. The major constituent of elastic fibres is elastin (Williams et al 1997).
fig.5. Elastic fibre (Taken from www.google.com)
The elastin arrangement varies depending largely on the strength and direction of forces on the tissue. Elastic fibres consist of an elastic core with some microfibrils located mostly around the periphery (fig 6). The exact mechanism of extensibility is not clearly understood. However, the quantity of elastin found within the tissue is usually reflected by the amount of mechanical strain inflicted on it and the requirement for reversible deformation. Some tissue, such as tracheobronhial tree of the lung, walls of arteries, and dermis are rich in elastic fibres, but some others such as ligaments, contain relatively sparse amount of elastic fibres, except the ligamenta nuchae in cervical spine and ligamenta flava of the thoracic and lumbar spine (Culav et al 1999).
Biosynthesis of elastin
Elastin is synthesized by smooth muscle cells and to a lesser extent by fibroblasts, but how it is synthesized is unclear. After elastin is secreted, it undergoes extensive cross-linking reactions with other elastin monomers and creates an extended insoluble fabric that provides tissue with elastic properties (Williams et al 1997).
A major problem in adults is the lack of regeneration of functional elastic fibres. This results in the inability to restore normal function. Elastin, however, is synthesized by adult tissue in response to cyclic stretching, injury, and ultraviolet radiation. A number of disease states, including emphysema, also lead to synthesis of elastin. Adults, apparently, cannot rebuild the elastic fibre assembly mechanism, and function is not restored. Generally, the mechanisms of control of elastic fibres formation are unknown (Culav et al 1999).
fig.6. Composition of elastic fibres
2. Proteoglycans (PGs)
According to their composition in type of glycosaminoglycan chain (GAG), all proteoglycans are named: chondroitin sulphates 4 (CS A) and 6 (CS C), keratan sulphate (KS), dermatan sulphate (DS), heparan sulphate (HS), and hyaluronan (HA) (Williams 1995, pages 84-86). They hydrate, stabilize and fill the space of the extracellular matrix (Culav et al 1999).
The PGs are consisted of a core protein at which are covalently attached one or more sulphated glycoasaminoglycan (GAG) chain, except hyaluronan, which neither attached to a protein core, nor sulphated (Hukins 1990, Williams 1995). However, it is usually included into the PG group, because it is the most abundant of the GAGs, and it has an important role in connecting to the other PGs to form supramolecular compound structures (Culav et al 1999).
There are various GAG chains with generally specific cores to each of the PG types. The GAG chains consist of repeating disaccharide units. The type and the number of units largely define the properties of the PG. Because of their negative polarization, all GAGs have a propensity to attract ions, and create a high osmotic activity that results in the absorption of water from surrounding tissues (Hukins 1990). The amount of hydration depends on the number of GAGs chains (Culav et al 1999). According to Williams et al (1997), the polyanionic character of the proteoglycans and the resulting swelling pressure keep the collagen fibrils apart and award stiffness on the porous gel, which they create.
The percentage of GAG with in CT varies directly proportional to the mechanical load. The higher the compressive forces acting on the tissue, the larger content of PG in it (approximately 8%-10% of the dry weight of the tissue). Conversely, in tissues, which are exposed in tension loads such as tendons and ligaments, PGs are concentrated in relatively small proportion (approximately 0,2% of dry weight). (Culav et al 1999).
fig.7. Very large proteoglycan (Taken from www.google.com)
According to Gray’s Anatomy (Williams 1995, page 86), the proteoglycans are classified into:
• Small PGs, which consist of the globular protein as the head and one or two glycan chain as the tails.
• Large PGs, which contain one or two protein cores attached by 5-10 glycan chains and
• Very large PGs, which are very complex molecules. They are consisted of 3 globular protein cores, which are bonded to hyaluronan. About 100 glycan chains are attached to each core (fig.7).
Culav et al (1999) divide the PGs in aggregating (large and very large) and nonaggregating (small) proteoglycans.
The small or nonaggregating PGs are attached at specific binding sites on the surface of collagen fibres and interact with them. The role of PGs in withstanding compression is limited, but they contribute to mechanical stability by their interaction with the collagen fibres. They are found in cartilage, disc, tendon, skin sclera, cornea.
The aggregating (large and very large) PGs are like the bristles of a bottlebrush, which fill the interfibrillar space. They are found in skin, sclera and tendon, and the very large ones are abundant in cartilage, disc and blood vessels. The large density of the aggregating PGs in cartilage and in the nucleus polposus of the intervertebral disc induces the movement of water into the matrix, resulting in the ability of these tissues to resist compressive forces (Culav et al 1999).
All PGs do not permit the neighbouring collagen fibres to been fused.
3. Glycoproteins.
Glycoproteins do not have mechanical functions, but they are important in stabilizing the surrounding matrix and linking the matrix to the cells. They regulate many functions of the cells such as producing changes in their shape, increasing their mobility and stimulating their proliferation. Among the best-known glycoproteins are fibronectin, tenascin, laminin, link protein, thrombospondin, osteropontin and fibromodulin.
Vessels and nerves of connective tissue
Generally, the blood supply of the connective tissue is poor, although some of the forms of the dense tissue, such as the dura mater and periosteum, have very rich blood support. Many blood vessels may pass through connective tissue en route for other tissues without supplying it (Williams 1995).
The lymphatic vessels are abundant in most types of the connective tissues, particularly beneath the skin. They are numerous in tendons, especially in their sheaths (Williams 1995).
Many kinds of nerve endings terminate in connective tissue and supply sensory innervation, which identify mechanical stresses, painful stimuli and thermal changes. Sympathetic innervation of the adipose tissue is important in controlling the metabolism of adipose cells, especially the breakdown of lipid (Williams 1995).
Human development begins with fertilization of an ovum. But between this initial contact and birth a series of complicated changes occurs. This involves both differentiation and reorganization of cells to form the different tissues, organs and organ systems to create a viable individual.
Immediately after fertilization the zygote undergoes repeated mitotic divisions, resulting in a ball of cells. When the cell has divided into a mass of sixteen cells (Now called the Morula) it enters the uterus wall. Differentiation continues so cells within the Morula are labeled depending on their position and function.
Simplification of the cell structure can label these into three zones: The Ectoderm, Endoderm and Mesoderm (Palastanga et al 1994).
The Ectoderm gives rise to the skin and its derivatives and the central and peripheral nervous system. The Endoderm forms many epithelial tissues of the body which lines many major bodily systems, e.g. Digestive and respiratory tracts. The Mesoderm forms various structures including the dermis, circulatory system, bone and cartilage.
The Mesoderm also gives rise to many specialist cells including Mesenchymal cells. These cells differentiate and specialize again to form the soft tissues, bones, muscles and nerves of the body.
Adapted from Gray’s (1995), these structures of the neuromusculoskeletal system can be divided into three distinct subgroups of Mesenchymal cells:
Firstly paraxial mesenchymal cells are responsible for the development of the vertebrae, portions of the neurocranium and axial skeleton. All voluntary muscles throughout the head, trunk, limbs, and the dermis of the skin.
The neural crest mesenchymal cells are responsible for the development of many soft tissues throughout the head including nerves, viscera, ligaments, tendons, fascia, muscular and dermal connectives tissue and the meninges. The neural crest also patterns the development of the nerves, muscles and blood vessels in this location. Schwann cells too, which are key regulators of peripheral nerve development originate from the neural crest and differentiate into myelin and non-myelin forming cells.
Finally the somatopleuric mesenchymal cells give rise to the skeletal elements, ligaments, tendons, fascia, muscular and dermal connective tissue throughout the trunk and limbs. These specific mesenchymal cells also pattern the development of nerves, muscles and blood vessels in these locations.
Summary
Human cells undergo specific differentiation and specialization immediately after fertilization. Basic cell structure can be split into three zones; ectoderm, endoderm and mesoderm. Connective tissues arise from specialist cells within the mesoderm zone. These mesenchymal cells are further divided into three distinct subgroups; paraxial, neural crest and somatopleuric. All connective tissues found within the body arise from neural crest and somatopleuric cells.
Cells Differentiation
Movement of cells populations (morphogenesis) and differentiation involves cells and tissue interactions in the early embryo. The process is known as (Gray 1995)
induction.
It is a complicated stage where the induced tissue may change or both tissues involved also may change in various combinations.
In Human Function and Structure,(Luciano 1978) cells, in fact, multiply, migrate and specialize, forming layers which can fold, fuse, separate and elongate in order to shape the human body. Other cells, which have not undergone cell differentiation, are known as stem cells. Some other undifferentiated cells can remain and multiply in the various organs where, under an appropriate stimulus, may then go through a delayed differentiation into a specialized cell type.
At the stage of fertilization every cell in the growing embryo receives an identical set of genes some of which are either turned off or on (i.e., from messenger RNA and the corresponding protein). Cell differentiation depends on the above mechanism suggesting a quantitative as well as qualitative control of gene expression.
Two types of interactions are described (Gray 1995): Permissive and Instructive; in a permissive interaction the signal between the apposing tissue and the responding tissue does not influence the developmental pathway selected. The responding tissue has the intrinsic capacity to develop and to express this capacity it only needs the appropriate environmental conditions.
Permissive interactions are often more common in a later stage of the development where the pre-determined fate of some tissue is only maintained and stabilized by another.
An instructive interaction changes the cell type of the responding tissue.
Condensation, (Hall BK 1992) is described as the first cellular product following tissue interaction and they are attributed a pivotal role in initiation of the vertebrate skeleton in embryonic development and in the modification of skeletal morphology during evolution.
Therefore, particularly important are the interactions between Epithelial and Mesenchymal cells during embryonic development, which take place in a reciprocal manner. These spatial, temporal sequential interactions both permissive and instructive leading to the differentiation and morphogenesis of most tissues and organs are termed epigenetic cascades.
It is recognized (Rogers 1983) that different signaling patterns take part during cells differentiation, influenced by size, shape, spacing, orientation and activity of many thousands of cells, each behaving in a reproducible way that is appropriate to its position in the tissue or organ concerned. The simplest way in which one cell can influence another is the release of some signaling molecule into the extracellular fluid. However, there are other ways to make sure that communication is transmitted and the following classification is an example:
Direct cell-cell contact;
Cell adhesion molecules;
Extracellular matrix and their receptors;
Growth factors and their receptors;
Ultimately, Connective Tissue (CT) is therefore the result of specific processes that occur during early embryonic development (Watkins 1999).
Tendons, ligaments and aponeurosis for example are the result and a combination of few cells and large amount of non -cellular material named matrix, produced by the cells. It is, in fact, the matrix the element that gives the various types of CT a different structure and function also.
Generally speaking CT has two functions:
Mechanical support, concerned with providing strength or elasticity as in binding together the cells of the body, supporting various organs and holding them into place, providing shock absorption and flexible links between bones and transmitting muscle forces.
Intercellular exchange, concerned with the circulation of body fluids, hence homeostasis.